

34. Хлоропласты (греч. Chloros — зеленый) — основная форма пластид. Они осуществляют фотосинтез. У растений хлоропласты содержатся не только в мезофилле, но и в клетках коровой паренхимы, флоэмы.
Морфология и ультраструктура хлоропластов. Форма, число, расположение в клетке хлоропластов весьма разнообразны. У зеленых водорослей может быть один хлоропласт на клетку, у эвгленовых и динофлагеллят молодые клетки содержат от 50 до 80 хлоропластов, старые — 200—300. Хлоропласты водорослей могут быть чашевидными, лентовидными, спиралевидными, пластинчатыми, звездчатыми и т.д.
В клетках разных растений содержится от 10 до 30 хлоропластов. В гигантских клетках палисадной ткани махорки их обнаружено около тысячи.
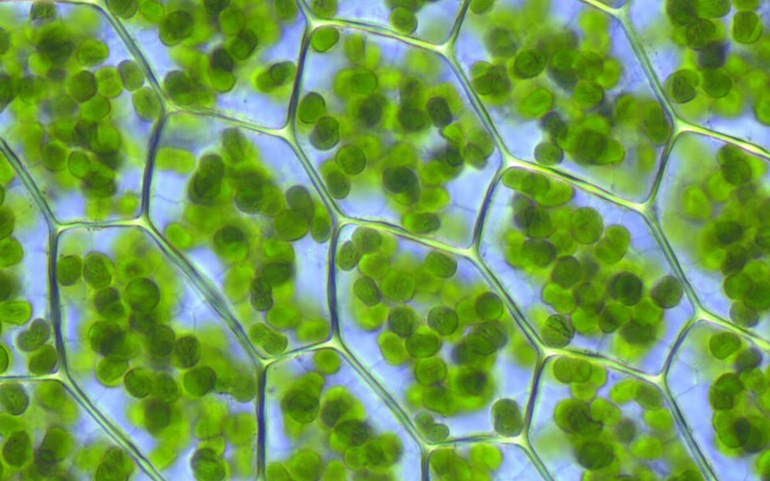
Хлоропласты растений представляют собой линзовидные образования, ширина которых составляет 2 — 4 мкм, длина — 5 мкм и более. Хлоропласт окружен двумя мембранами, которые разделены узким межмембранным (перипластидным) пространством шириной около 20 — 30 нм. Наружная мембрана обладает высокой проницаемостью, внутренняя — менее проницаема и несет специальные транспортные белки. Внутренняя мембрана окружает большую центральную область — строму.
Строма хлоропласта содержит разнообразные ферменты, рибосомы, ДНК и РНК. В строме хлоропластов располагаются небольшие мембранные мешочки – тилакоиды. Мембрана тилакоидов включает в свой состав пигмент-белковые комплексы, прежде всего основной пигмент растений — хлорофилл, существующий в нескольких формах, каротиноиды, из которых наиболее обычны каротины и ксантофиллы. Кроме того, в тилакоидную мембрану включены компоненты электрон-транспортных цепей. Внутренние полости тилакоидов создают внутренний компартмент хлоропласта — тилакоидное пространство. Тилакоиды образуют стопки — граны, в одной стопке от нескольких штук до 50 и более тилакоидов. Размер гран в зависимости от числа тилакоидов в них может достигать 0,5 мкм, в этом случае они доступны для наблюдений с помощью светового микроскопа. На один хлоропласт приходится 40 — 60 гран. Тилакоиды в гранах располагаются на расстоянии около 20 нм. В состав гран кроме тилакоидов входят участки тилакоидов стромы.
У водорослей кроме хлорофилла и каротиноидов присутствуют: фикоцианин, фикоэритрин, фукоксантин, часто маскирующие основную зеленую окраску.
Еще одна особенность строения хлоропластов водорослей, — присутствие в них пиреноидов (греч. pyrenoideus — имеющий вид косточки). Это плотное образование белковой природы, вокруг которого концентрируется крахмал. Пиреноиды характерны для хлоропластов зеленых водорослей.
Функция хлоропластов — фотосинтез, образование органических веществ из диоксида углерода и воды за счет энергии солнечного света. Разнообразные реакции, протекающие при фотосинтезе, можно разделить на две большие группы. Первая — это реакции фотосинтетического переноса электронов, или световые реакции. Вторая — реакции фиксации углерода, или темповые реакции. Световые реакции осуществляются в тилакоидах. Темновые реакции происходят в строме хлоропласта.
Количество хлоропластов в клетке поддерживается двумя способами: путем деления и за счет преобразования лейкопластов. В ходе этого процесса усложняется внутренняя ламеллярная структура пластиды, возникают граны, увеличиваются их число и размер. Одновременно увеличиваются размеры пластиды.
Лейкопласты (греч. leucos — белый) — бесцветные пластиды, содержащиеся в клетках органов растений, лишенных окраски. Часто это запасающие органы и ткани. Лейкопласты представляют собой округлые образования, наибольший размер которых составляет 2 — 4 мкм. Они окружены оболочкой, состоящей из двух мембран, внутри которой находится белковая строма. Строма лейкопластов содержит небольшое число пузырьков и плоских цистерн — ламелл. Основная функция лейкопластов – вторичный синтез веществ.
Лейкопласты способны превращаться в хлоропласты, процесс их развития связан с увеличением размеров, усложнением внутренней структуры и образованием зеленого пигмента — хлорофилла. Такая перестройка пластид происходит, например, при позеленении клубней картофеля. Лейкопласты способны также переходить в хромопласты, например в корнеплоде моркови. В некоторых тканях, таких, как эндосперм зерновки злаков, в корневищах и клубнях лейкопласты превращаются в хранилище запасного крахмала — амилопласты. Гранулы крахмала полностью заполняют всю его полость.
Процесс развития различных пластид представляет собой монотропно направленный ряд, т.е. идущий в одном направлении:
лейкопласт → хлоропласт → хромопласт
Онтогенетические переходы одной формы в другую необратимы; хромопласт не может сформировать ни хлоропласта, ни лейкопласта. Точно так же хлоропласт не может вернуться в состояние лейкопласта.
Хромопласты — желто-оранжевые пластиды. Они имеют овальную, ромбовидную или заостренную в виде полумесяца форму. Образуются из хлоропластов, при деградации последних, реже – из лейкопластов, минуя стадию зеленых пластид, как в корне моркови.
Роль хромопластов в клетке неясна. Но для растительных организмов в целом эти пластиды играют важную биологическую роль. Органы растений, в которых прекращаются процессы фотосинтеза, становятся привлекательными для насекомых, птиц, других животных, которые опыляют растения и распространяют плоды и семена. При осеннем пожелтении листьев разрушение хлоропластов и образование хромопластов приводит к утилизации белков и азотсодержащих соединений, которые перед листопадом оттекают в другие органы растения.
36. Пластиды (греч. plastides — созидающие, образующие) — это двумембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов — растений, одноклеточных и многоклеточных водорослей. У растений существует комплекс различных пластид, отличающихся строением и функциями.
Всем пластидам свойствен ряд общих черт. Они имеют свой генетический аппарат и окружены оболочкой, состоящей из двух элементарных мембран. Все пластиды развиваются из пропластид. Пропластиды представляют собой мелкие органоиды, присутствующие в клетках меристемы, судьба которых определяется потребностями дифференцированных клеток.
Выделяют три основных типа пластид: лейкопласты, хлоропласты и хромопласты. Все они представляют собой генетически связанный ряд.
Происхождение пластид. Биохимические данные дают основания считать, что хлоропласты — это потомки фотосинтезирующих цианобактерий, ко¬торые были захвачены путем эндоцитоза свободноживущими эукариотическими клетками и перешли к симбиозу с ними. Генетический аппарат хлоропластов и аппарат белкового син¬теза хорошо изучены как у растений, так и у водорослей. Следует подчеркнуть, что гены хлоропластов у растений, имею¬щих даже отдаленное родство, практически одинаковы и обнару¬живают большое сходство с бактериальным геномом. Это рассмат-ривается как одно из убедительных доказательств теории эндосимбионтов. В частности, ДНК хлоропластов кольцевая, как у бак¬терий, и включает около 120 генов, что существенно меньше, чем в бактериальной клетке. Белки, кодируемые в хлоропластах, очень похожи на бактериальные, а некоторые группы генов с близ¬кими функциями организованы одинаково как в хлоропластах растений, так и в кишечной палочке, и у цианобактерий. Анализ нуклеотидного состава ДНК хлоропластов печеночного мха и табака показал их почти полное сходство. По-видимому, геном хлоропластов остается почти неизменным по меньшей мере несколько сотен миллионов лет, именно столько лет назад разо¬шлись эволюционные пути этих ветвей живых организмов. Белоксинтезирующий аппарат хлоропластов также близок к бактериальному. Рибосомы пластид типа 70 S, как у бактерий, и, более того, рибосомы хлоропластов могут использовать тРНК бак¬терий при синтезе белка.
38. Ядро клетки – это место хранения, воспроизведения и начальной реализации наследственной информации в эукариотической клетке.
Впервые ядро клетки зарисовал Ф. Фонтана (1781 г.), затем Я. Пуркинье (1825 г.), но только Р. Браун (1831-1833 гг.) дал ему название nucleus и доказал, что ядро имеется во всех растительных и животных клетках. В 1833 г. Р. Броун опубликовал результаты своих микроскопических наблюдений над орхидеями. В их клетках он обнаружил и описал постоянные шаровидные структуры, которые обозначил термином «ядро» (греч. karion — ядро, лат. nucleus — ядро). Позднее ядра были найдены во всех эукариотических клетках. Объем ядра составляет около 10 % общего объема клетки.
Ядро может иметь различную форму: округлую, эллипсоидальную, продолговатую (палочковидную, нитевидную), сегментированную. Например, в зернистых лейкоцитах ядро подразделяется на сегменты (сегментоядерные лейкоциты).
Как правило, в клетке имеется одно ядро. Однако известны многоядерные клетки: например, инфузории-туфельки имеют два ядра (макро- и микронуклеус), а в клетках других низших эукариот может содержаться несколько десятков и сотен ядер. Локализуется ядро обычно в центре клетки, но в клетках эпителиальных тканей ядра нередко сдвинуты к базальному полюсу.
В клетках прокариот (эубактерии, цианобактерии, архебактерии) оформленного ядра нет. Его функции выполняет нуклеоид, основу которого составляет бактериальная хромосома. Дополнительными носителями генетической информации у прокариот являются плазмиды – мелкие кольцевые молекулы ДНК.
Некоторые специализированные клетки лишены ядра. Например, ситовидные трубки растений, зрелые эритроциты млекопитающих, тромбоциты, роговые чешуйки, являются вторичными (постклеточными) образованиями, так как они образуются из ядерных клеток в результате их специфической дифференцировки. Клетки, искусственно лишенные ядра, называются энуклеированными. Энуклеированные клетки утрачивают способность к делению, а продолжительность их жизни резко сокращается.
Хлоропласты – особенности строения, функции и роль в фотосинтезе
Функциональные особенности
Строение хлоропласта изучается школьниками в 6 классе на уроках биологии. К особенностям клеток относится наличие в строме рибосомы, ДНК, РНК. В мембране присутствует вещество, способное придать растениям соответствующий цвет. Для хлорофилла характерен зеленый оттенок, а для каротиноида:

Значение хлорофилла для растений заключается в возможности осуществления процесса фотосинтеза. С учётом строения биологи выделяют 4 типа хлорофилла: a, b, c, d. Первые два содержатся в растениях на суше и зеленых водорослях. Типы a и c считаются растительными компонентами диатомовых, d и a — красных водорослей.
Для хлорофилла характерно поглощение солнечной энергии с последующей передачей иным молекулам. Разрушение зеленого вещества наблюдается в конце жизненного цикла органоида в результате резкого изменения светового дня и значения температуры. Часть хлоропластов превращается в хромопласты. Это приводит к изменению внутренней информации, появлению нового цветового оттенка, опадению листьев.
Принципы классификации
Пластиды делятся на три вида: лейкопласты (бесцветные), хлоропласты (окрашенные в зеленый цвет), хромопласты (имеют разные оттенки). На протяжении жизни клетки способны превращаться друг в друга. Лейкопластам свойственно переходить в хлоропласты, а последние за счёт появления бурых и прочих пигментов — в хромопласты, пластоглобулы.
Внешне зеленые вещества покрыты липидной и белковой мембранами. Полужидкая строма с тилакоидами (компартменты, ограниченные мембраной) считается основным веществом, в состав которого входят граны с каналами. Первые компоненты представлены в виде плоских круглых мешочков, расположенных перпендикулярно поверхности двухмембранных органоидов (ДО).
Уникальность их структуры заключается в хранении зеленого пигмента (хлорофилл). Главная функция хлоропластов связана с участием в фотосинтетическом явлении. В их состав входят жиры, зерна (митохондрия, пропластида), крахмал.
На долю липидов приходится до 30%. Они представлены тремя группами:
- Структурная. В состав входят амфипатические вещества.
- Гидрофобная. В группу входят каротиноиды, которые защищают зеленые вещества от фотоокисления. Одновременно они транспортируют водород.
- Жирорастворимая. Группа состоит из витаминов К и Е.
К другим компонентам, входящим в состав хлоропласта, относятся углеводы. Они представлены в виде продуктов фотосинтеза. До 25% приходится на долю минералов. Ферменты могут выполнять двойную функцию: катализацию различных реакций, обеспечение биосинтеза белков.
Внутренняя структурированность хлоропластов зависит от функциональных нагрузок, физиологического состояния. Молодые клетки размножаются за счет деления, а зрелые обладают выраженной системой гран. Если они стареют, происходит разрыв тилакоидов, распадается хлорофилл. Осенью деградация приводит к появлению хромопластов.
Главная роль хлоропластов в фотосинтезе обеспечена их способностью пассивно двигаться в клетках, увлекаемых током цитоплазмы. Веществу свойственно собирать свет и активно перемещаться с одного места на другое. При интенсивном свете оно поворачивается ребром к яркому солнцу, выстраиваясь вдоль стенок, которые параллельны лучам.
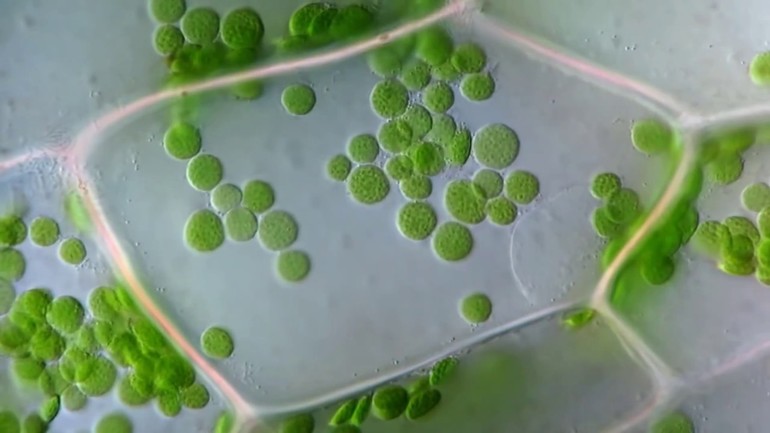
Если освещение слабое, схема движения хлоропластов следующая: они перемещаются на стенки, обращённые к солнцу, поворачиваясь наибольшей поверхностью. Когда освещение среднее, клетки занимают соответствующее положение. От условий освещения зависит то, какие пигменты хлоропластов появятся.
Для пластид и митохондрий свойственна полуавтономная степень. Кроме фотосинтеза, в первых компонентах происходит биосинтез белка. Так как они содержат в себе ДНК, поэтому принимают активное участие в наследственном комплексе: передача признаков, цитоплазматические свойства.
Описание хромопластов
К пластидам высших растений относятся хромопласты. Они имеют незначительные размеры. Для внутриклеточных органелл характерен разный окрас: красный, желтый, коричневый. Он придает соответствующий цвет осенью, плодам и цветкам, что необходимо для привлечения опылителей и животных, разносящих семена продолжительные расстояния.
Структура ткани похожа на иные пластиды. Внутренняя оболочка развита слабее внешней. У некоторых представителей она может отсутствовать. В каротиноидах (жирорастворимые пигменты) происходит накапливание кристаллов. Для определения точных функций вещества изучается таблица с формами хромопластов:

Их роль в жизни растений до конца не выяснена. Ученые предполагают, что пигменты участвуют в окислительных и восстановительных процессах, необходимых для размножения и физиологического развития клеток.
Строение лейкопластов
В органоидах этого типа накапливаются питательные компоненты. Лейкопласты имеют 2 оболочки: внутреннюю и внешнюю. На свету им свойственно превращаться в хлоропласты, но в привычном состоянии органоиды бесцветны. Основная их форма — шаровидная. Размещены они в мягких частях растений:
С учетом накапливаемого вещества лейкопласты классифицируются на следующие виды: амилопласты, элайопласты, протеинопласты. В первую группу входят органоиды с крахмалом, находящиеся в каждом растении. Если лейкопласт полностью заполнен крахмалом, он называется крахмальным зерном. Для элайопластов характерно продуцирование и запас жиров, а для протеинопластов — скопление белковых веществ.
Лейкопласты обладают ферментной субстанцией, что способствует ускоренному протеканию химических реакций. В отрицательном жизненном периоде, когда не происходит фотосинтез, они расщепляют полисахариды на простые углеводы. Так как в луковицах содержится много органоидов, поэтому им свойственно переносить длительную засуху, жару, низкую температуру. После выполнения своих функций они становятся хромопластами.
Симбиотическая теория
Чтобы выяснить механизм появления пластид, митохондрий и других органоидов, рассматривается теория эндосимбиоза. Ее суть заключается в совместной и взаимовыгодной жизни органеллы с клеткой. Впервые теорию предложил Шимпер в 1883 году. В 1867 ученые работали над двойственной природой лишайников.
Биолог Фамицын, учитывая теорию Шимпера, предположил, что хлоропласты, как лишайники и водоросли, относятся к симбионтам. Ученые доказали, что митохондрии — аэробные бактерии, которые не размножаются за пределами клеток. Общие свойства, характерные для митохондрий и пластид:
- наличие двух замкнутых мембран;
- размножение бинарным делением;
- ДНК не связана с гистонами;
- наличие своего аппарата синтеза белка.
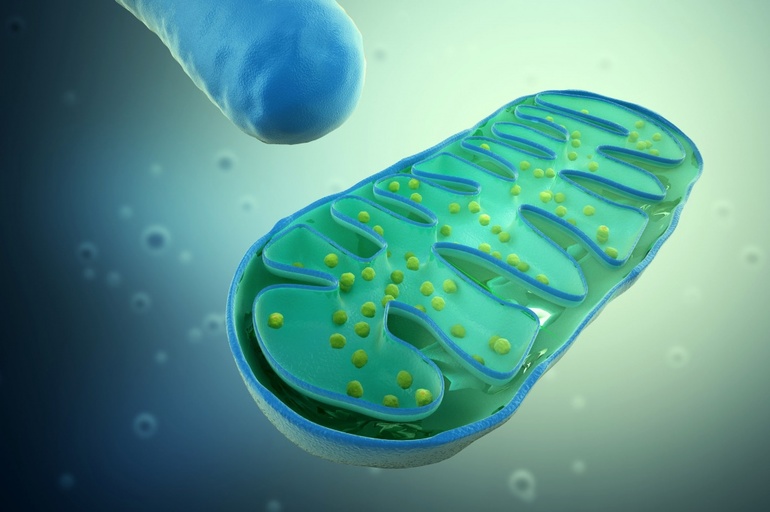
В ДНК пластид и митохондрий, в отличие от аналогичных структур прокариот, нет интронов. А в ДНК хлоропластов закодирована информация о некоторых белках, остальные данные находятся в ядре клетки. В результате эволюции часть генетического материала из генома перешло в ядро, поэтому хлоропласты и митохондрии не размножаются независимо.
Археи и бактерии не склонны к фагоцитозу. Они питаются только осмотрофно. Множественные биологические и химические исследования указывают на химерную сущность бактерий. Ученые не выяснили, как сливаются организмы из нескольких доменов. В условиях современности выявлены организмы, которые содержат в себе другие клетки в качестве эндосимбионтов. Они отличаются от первичных эукариотов тем, что не интегрируются в одно целое, не имеют своей индивидуальности.
Интересным организмом считается Mixotricha paradoxa. Чтобы двигаться, она использует 250 000 бактерий, которые фиксируются на ее поверхности. Митохондрии у этого организма вторично потеряны. Внутри находятся сферические аэробные микроорганизмы, которые заменяют органеллы.
Пластиды: виды, строение и функции. Хлоропласты, хромопласты, лейкопласты
Пластиды — органоиды, специфичные для клеток растений (они имеются в клетках всех растений, за исключением большинства бактерий, грибов и некоторых водорослей).
В клетках высших растений находится обычно от 10 до 200 пластид размером 3-10мкм, чаще всего имеющих форму двояковыпуклой линзы. У водорослей зеленые пластиды, называемые хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую, лентовидную, сетчатую и другие формы.
Различают 3 вида пластид:
- Бесцветные пластиды — лейкопласты;
- окрашенные — хлоропласты (зеленого цвета);
- окрашенные — хромопласты (желтого, красного и других цветов).
Эти виды пластид до известной степени способны превращаться друг в друга — лейкопласты при накоплении хлорофилла переходят в хлоропласты, а последние при появлении красных, бурых и других пигментов — в хромопласты.
Строение и функции хлоропластов
Хлоропласты — зеленые пластиды, содержащие зеленый пигмент — хлорофилл.
Основная функция хлоропласт — фотосинтез.
В хлоропластах есть свои рибосомы, ДНК, РНК, включения жира, зерна крахмала. Снаружи хлоропласта покрыты двумя белково-липидными мембранами, а в их полужидкую строму (основное вещество) погружены мелкие тельца — граны и мембранные каналы.
 Строение хлоропласта
Строение хлоропласта
Граны (размером около 1мкм) — пакеты круглых плоских мешочков (тилакоидов), сложенных подобно столбику монет. Располагаются они перпендикулярно поверхности хлоропласта. Тилакоиды соседних гран соединены между собой мембранными каналами, образуя единую систему. Число гран в хлоропластах различно. Например, в клетках шпината каждый хлоропласт содержит 40-60 гран.
Хлоропласты внутри клетки могут двигаться пассивно, увлекаемые током цитоплазмы, либо активно перемещаться с места на место.
- Если свет очень интенсивен, они поворачиваются ребром к ярким лучам солнца и выстраиваются вдоль стенок, параллельных свету.
- При слабом освещении, хлоропласты перемещаются на стенки клетки, обращенные к свету, и поворачиваются к нему своей большой поверхностью.
- При средней освещенности они занимают среднее положение.
Этим достигаются наиболее благоприятные для процесса фотосинтеза условия освещения.
Хлорофилл
В гранах пластид растительной клетки содержится хлорофилл, упакованный с белковыми и фосфолипидными молекулами так, чтобы обеспечить способность улавливать световую энергию.
Молекула хлорофилла очень сходна с молекулой гемоглобина и отличается главным образом тем, что расположенный в центре молекулы гемоглобина атом железа заменен в хлорофилле на атом магния.
 Сходство молекулы хлорофилла и молекулы гемоглобина
Сходство молекулы хлорофилла и молекулы гемоглобина
В природе встречается четыре типа хлорофилла: a, b, c, d.
Хлорофиллы a и b содержат высшие растения и зеленые водоросли, диатомовые водоросли содержат a и c, красные — a и d.
Лучше других изучены хлорофиллы a и b (их впервые разделил русский ученый М.С.Цвет в начале XXв.). Кроме них существуют четыре вида бактериохлорофиллов — зеленых пигментов пурпурных и зеленых бактерий: a, b, c, d.
Большинство фотосинтезирующих бактерий содержат бактериохлорофилл a, некоторые — бактериохлорофилл b, зеленые бактерии — c и d.
Хлорофилл обладает способностью очень эффективно поглощать солнечную энергию и передавать ее другим молекулам, что является его главной функцией. Благодаря этой способности хлорофилл — единственная структура на Земле, которая обеспечивает процесс фотосинтеза.
Главная функция хлорофилла в растениях — поглощение энергии света и передача ее другим клеткам.
Пластидам, так же, как и митохондриям, свойственна до некоторой степени автономность внутри клетки. Они размножаются путем деления.
Наряду с фотосинтезом, в пластидах происходит процесс биосинтеза белка. Благодаря содержанию ДНК пластиды играют определенную роль в передаче признаков по наследству (цитоплазматическая наследственность).
Строение и функции хромопластов
Хромопласты относятся к одному из трех видов пластид высших растений. Это небольших размеров, внутриклеточные органеллы.
Хромопласты имеют различный окрас: желтый, красный, коричневый. Они придают характерный цвет созревшим плодам, цветкам, осенней листве. Это необходимо для привлечения насекомых-опылителей и животных, которые питаются плодами и разносят семена на дальние расстояния.
 Строение хромопласта
Строение хромопласта
Структура хромопласта похожа на другие пластиды. Их двух оболочек внутренняя развита слабо, иногда вовсе отсутствует. В ограниченном пространстве расположена белковая строма, ДНК и пигментные вещества (каротиноиды).
Каротиноиды – это жирорастворимые пигменты, которые накапливаются в виде кристаллов.
Форма хромопластов очень разнообразна: овальная, многоугольная, игольчатая, серповидная.
Роль хромопластов в жизни растительной клетки до конца не выяснена. Исследователи предполагают, что пигментные вещества играют важную роль в окислительно-восстановительных процессах, необходимы для размножения и физиологичного развития клетки.
Строение и функции лейкопластов
Лейкопласты — это органоиды клетки, в которых накапливаются питательные вещества. Органеллы имеют две оболочки: гладкую наружную и внутреннюю с несколькими выступами.
Лейкопласты на свету превращаются в хлоропласты (к примеру зеленые клубни картофеля), в обычном состоянии они бесцветны.
Форма лейкопластов шаровидная, правильная. Они находятся в запасающей ткани растений, которая заполняет мягкие части: сердцевину стебля, корня, луковиц, листьев.
 Строение лейкопласта
Строение лейкопласта
Функции лейкопластов зависят от их вида (в зависимости от накапливаемого питательного вещества).
- Амилопласты накапливают крахмал, встречаются во всех растениях, так как углеводы основной продукт питания растительной клетки. Некоторые лейкопласты полностью наполнены крахмалом, их называют крахмальными зернами.
- Элайопласты продуцируют и запасают жиры.
- Протеинопласты содержат белковые вещества.
Лейкопласты также служат ферментной субстанцией. Под действием ферментов быстрее протекают химические реакции. А в неблагоприятный жизненный период, когда процессы фотосинтеза не осуществляются, они расщепляют полисахариды до простых углеводов, которые необходимы растениям для выживания.
В лейкопластах не может происходить фотосинтез, потому что они не содержат гран и пигментов.
Луковицы растений, в которых содержится много лейкопластов, могут переносить длительные периоды засухи, низкую температуру, жару. Это связано с большими запасами воды и питательных веществ в органеллах.
Предшественниками всех пластид является пропластиды, небольшие органоиды. Допускают, что лейко — и хлоропласты способны трансформироваться в другие виды. В конечном итоге после выполнения своих функций хлоропласты и лейкопласты становятся хромопластами — это последняя стадия развития пластид.
Важно знать! Одновременно в клетке растения может находиться только один вид пластид.
Сводная таблица строения и функций пластид
| Свойства | Хлоропласты | Хромопласты | Лейкопласты |
|---|---|---|---|
| Строение | Двухмембранная органелла, с гранами и мембранными канальцами | Органелла с не развитой внутренней мембранной системой | Мелкие органеллы, находятся в частях растения, скрытых от света |
| Окрас | Зеленые | Разноцветные | Бесцветные |
| Пигмент | Хлорофилл | Каротиноид | Отсутствует |
| Форма | Округлая | Многоугольная | Шаровидная |
| Функции | Фотосинтез | Привлечение потенциальных распространителей растений | Запас питательных веществ |
| Заменимость | Переходят в хромопласты | Не изменяются, это последняя стадия развития пластид | Превращаются в хлоропласты и хромопласты |
Хлоропласты: роль в процессе фотосинтеза и структура
Фотосинтез происходит в эукариотических клеточных структурах, называемых хлоропластами. Хлоропласт – это тип органеллы растительных клеток, известный как зеленые пластиды. Пластиды помогают хранить и собирать необходимые вещества для производства энергии. Хлоропласт содержит зеленый пигмент, называемый хлорофиллом, который поглощает световую энергию для процесса фотосинтеза. Следовательно, название хлоропласт указывает на то, что эти органеллы представляют собой хлорофиллсодержащие пластиды.
Подобно митохондриям, хлоропласты имеют свою собственную ДНК, ответственны за производство энергии и воспроизводятся независимо от остальной части клетки посредством процесса деления, подобного бактериальному бинарному делению. Они также ответственны за производство аминокислот и липидных компонентов, необходимых для производства хлоропластов. Хлоропласты также встречаются в клетках других фотосинтезирующих организмах, таких как водоросли.
Хлоропласт: структура
Схема строения хлоропласт
Хлоропласты обычно встречаются в охранных клетках, расположенных в листьях растений. Охранные клетки окружают крошечные поры, называемые устьицами, открывая и закрывая их, чтобы обеспечить необходимый для фотосинтеза газообмен. Хлоропласты и другие пластиды развиваются из клеток, называемых пропластидами, которые являются незрелыми, недифференцированными клетками, развивающимися в разные типы пластид. Пропластид, развивающийся в хлоропласт, осуществляет этот процесс только при свете. Хлоропласты содержат несколько различных структур, каждая из которых имеет специализированные функции. Основные структуры хлоропласта включают:
- Мембрана – содержит внутренние и внешние липидные двухслойные оболочки, которые выступают в качестве защитных покрытий и сохраняют замкнутые структуры хлоропластов. Внутренняя мембрана отделяет строму от межмембранного пространства и регулирует прохождение молекул в/из хлоропласта.
- Межмембранное пространство – пространство между внешней и внутренней мембранами.
- Тилакоидная система – внутренняя система мембран, состоящая из сплющенных мешкообразных мембранных структур, называемых тилакоидами, которые служат местами преобразования энергии света в химическую энергию.
- Тилакоид с просветом (люменом) – отсек в каждом тилакоиде.
- Грана – плотные слоистые стопки тилакоидных мешков (10-20), которые служат местами преобразования энергии света в химическую энергию.
- Строма – плотная жидкость внутри хлоропласта, содержащая внутри оболочки, но вне тилакоидной мембраны. Здесь происходит конверсия углекислого газа в углеводы (сахара).
- Хлорофилл – зеленый фотосинтетический пигмент в хлоропласт-гране, поглощающий световую энергию.
Хлоропласт: фотосинтез
При фотосинтезе энергия солнечного света преобразуется в химическую энергию. Химическая энергия хранится в виде глюкозы (сахара). Двуокись углерода, вода и солнечный свет используются для производства глюкозы, кислорода и воды. Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Световая фаза фотосинтеза протекает только при наличии света и происходит внутри хлоропластовой граны. Первичным пигментом, используемым для преобразования световой энергии в химическую, является хлорофилл а. Другие пигменты, участвующие в поглощении света, включают хлорофилл b, ксантофилл и каротин. Во время световой фазы, солнечный свет преобразуется в химическую энергию в виде АТФ (молекулы, содержащей свободную энергию) и НАДФ (молекула, несущая электроны высокой энергии).
И АТФ, и НАДФ используются во время темновой фазы для получения сахара. Темновая фаза фотосинтеза, также известная как этап фиксации углерода или цикл Кальвина. Реакции на этой стадии возникают в строме. Строма содержит ферменты, которые облегчают серию реакций, использующих АТФ, НАДФ и углекислый газ для получения сахара. Сахар может храниться в виде крахмала, используемого во время дыхания или при производстве целлюлозы.
Экология СПРАВОЧНИК
Информация
Добавить в ЗАКЛАДКИ
| Поделиться: | Пластиды ХлоропластыХлоропласты — внутриклеточные органеллы растительной клетки — зеленые пластиды, в которых осуществляется процесс фотосинтеза. Окрашены в зеленый цвет хлорофиллами, содержат также желтые пигменты — каротино-иды.[ . ] Хлоропласты (зеленые пластиды)—органеллы клетки, обусловливающие накопление углеводов в процессе фотосинтеза. В них содержится большое количество ферментов, контролирующих фотосинтез, а также белков, жирных кислот и фосфолипидов. Эти важнейшие мембранные органеллы встречаются в клетках эукариотических организмов: высших растений и некоторых одноклеточных.[ . ] Пластиды — специализированные дискообразные тела, содержащиеся в цитоплазме и свойственные только растительным клеткам. По наличию или отсутствию пигмента пластиды делятся на лейкопласты (бесцветные) и хромопласты (окрашенные). Лейкопласты присутствуют во взрослых клетках, не подвергающихся воздействию света, а некоторые типы включены в запасной крахмал. Из окрашенных наибольшее значение имеют те пластиды, которые содержат хлорофилл (хлоропласты), так как они являются структурно и функционально полными элементами фотосинтеза, процесса, в котором углекислый газ и вода превращаются в углеродсодержащие соединения. Эти реакции приводят к образованию крахмальных зерен в хлоропла-стах.[ . ] Для пластид характерны переходы из одного типа в другой. Например, зеленые хлоропласты листьев переходят в хромопласты (осенью, когда изменяется окраска листьев), а лейкопласты зеленеющих клубней картофеля переходят в хлоропласты.[ . ] Строение хлоропластов несколько напоминает строение митохондрий, Их тело (матрикс, или строма) ограничено двумя белково-липидными мембранами, толщиной 7 нм каждая. Наружный слой, соприкасающийся с цитоплазмой, обычно имеет ровные контуры и не образует выпячиваний или складок, внутренний, как и у других пластид, образует складчатые выпячивания внутрь стромы.[ . ] Для развития хлоропластов из пропластид обычно необходим свет. Развитие происходит путем отшнурсмвывания от внутренней мембраны уплощенных пузырьков, образующих сплющенную, окруженную двойной мембраной пластинку тилакоидов, в которых в конечном итоге синтезируется хлорофилл. Пластиды, которые развиваются таким образом, ■ называются этиопластами.[ . ] У водорослей хлоропласты могут иметь самую разнообразную форму и размеры, а у высших растений они очень однородны по своей форме. Эти пластиды окружены двойной мембраной и состоят из бесцветного жидкого матрикса (стромы) с находящейся в нем сложной системой уплощенных мембранных пузырьков, известных как тилакоиды, которые объединяются в стопки (граны). Тилакоиды имеют округлые очертания и соединяются с тилакоидами в другой гране посредством связующих мембран (рис. 1.8). Мембраны тилакоидов .состоят из-липидов и белков, к которым присоединяется хлорофилл.[ . ] Кроме зеленых хлоропластов, для каулерповых характерно наличие бесцветных пластид, или лейкопластов, которые участвуют в процессе накопления крахмала. У всех других водорослей порядка найдены только хлоропласты.[ . ] Различают 3 типа пластид: бесцветные — лейкопласты, зеленые — х л о р о-пласты, окрашенные в другие цвета — хромопласты. Пластиды каждого типа имеют свое строение и несут свои, им присущие функции. Однако возможны переходы пластид из одного типа в другой. Так, позеленение клубней картофеля вызывается перестройкой их лейкопластов в хлоропласты. В корнеплоде моркови лейкопласты переходят в хромопласты. Пластиды всех трех типов образуются из пропластид.[ . ] Таким образом, на примере хлоропласта видно, что ацетилхолин, находящийся в цитоплазме и строме пластиды, может вызвать открывание ионных каналов и уменьшение мембранного потенциала, который при распространении по мембране может представлять собой внутриклеточный ПД. Аналогичное явление, вызванное открыванием Са2+-каналов, при действии катехоламинов и серотонина также выявлено в хлоропластах.[ . ] Для выяснения локализации фермента в пластидах было проведено разделение хлоропластов на фракции в градиенте плотности сахарозы. Холинэстеразная активность была обнаружена во фракциях наружных мембран и тилакоидов. При этом активность холинэстеразы в тилакоидах была примерно в 7 раз выше, чем в оболочке хлоропластов.[ . ] В сформировавшейся клетке растущих листьев пластиды размножаются путем деления. Деление может происходить или па стадии пропластид (у высших растений). или ва стадии уже сформировавшихся хлоропластов (у водорослей и папоротников). Пнастиды делятся или путем возникновения поперечных перегородок, или, реже, путем почкования. Поперечные перегородки образуются за счет образования складок на внутренней мембране. До тех пор, пока складка внутренней мембрапы ие поделила тело пластид, наружная мембрана сохраняется и объединяет обе дочерние пластиды.[ . ] Различают три вида пластид: лейкопласты — бесцветные, хромопласты — оранжевые, хлоропласты — зеленые. Именно в хлоро-пластах сосредоточен зеленый пигмент хлорофилл.[ . ] Современные исследователи (Матиенко, 1965) делят пластиды на две группы: лейкопласты (бесцветные пластиды) и хромопласты (пластиды, содержащие пигменты). В группу лейкопластов входят: амилопласты — пластиды, накапливающие крахмал; олеопласты — пластиды, накапливающие масла, и протеинопла-сты — пластиды, накапливающие белок. В группу хромопластов входят: хлоропласты — зеленые пластиды, в основном содержащие хлорофилл, и каротинопласты—пластиды, содержащие желтые пигменты.[ . ] Для растительной клетки характерно присутствие пластид. Важнейшие пластиды — это хлоропласты. Они осуществляют трансформацию световой энергии в химическую. Другой важнейший энергетический процесс (синтез АТФ за счет энергии окисления) происходит в митохондриях. Они представляют собой овальные или палочковидные структуры длиной 1. 2 мкм. Система канальцев и цистерн (диктиосом), ограниченных однослойной мембраной, составляет аппарат Гольджи, основная функция которого — внутриклеточная секреция веществ, необходимых для построения клеточной оболочки и др. В округлых тельцах — лизосомах сконцентрированы гидролитические ферменты. С помощью сферосом идет синтез липидов.[ . ] В клетках растений вместо митохондрии содержатся пластиды. Среди пластид различают хлоропласты, которые содержат хлорофилл, лейкопласты (бесцветные пластиды), в которых происходит накопление крахмала, и хромопласты, в которых происходит синтез пигмента плодов. У пурпурных бактерий имеются хроматофоры.[ . ] В настоящее время получены данные, что гиббереллины содержатся в пластидах, где могут проходить по крайней мере некоторые, а быть может, и все стадии их биосинтеза. Следовательно, хлоропласты синтезируют свою собственную мевалоновую кислоту, часть которой используется для синтеза гиббереллииов. Могут ли и другие клеточные органеллы наряду с пластидами синтезировать гиббереллины, остается пока неясным.[ . ] Во время деления клетки происходит репликация различных органелл, в том числе пластид и митохондрий. Простейшим типом пластиды является пропластида, из которой развиваются все типы пластид, включая хлоропласта. Пластиды представляют собой полуавтономные оргаиеллы, способные к удвоению путем деления или почкования. В клетках высших растений может содержаться от нескольких до большого числа пластид, и клетки различных типов значительно отличаются друг от друга по содержанию в них пластид. Число пластид в клетках какого-либо одного типа обычно остается приблизительно постоянным, и это наводит на мысль, что репликация пластид происходит одновременно с делением клетки. Однако распределение пластид материнской клетки между дочерними происходит, по-видимому, случайным образом.[ . ] Возбудитель болезни — Bean common mosaic virus (Phaseo-lus virus 1 Smith), имеет ямочковидную форму. Он вызывает разрушение хлоропластов и утолщение стромы пластид. Вирус обыкновенной мозаики фасоли передается с семенами, во время вегетации растений — многими видами тлей, :à иногда инокуляцией сока. Выдерживает разведение 1:1000 и инактивируется при температуре 56—58 °С в течение 10 мин.[ . ] Основная функция хлородластов — это процесс фотосинтеза. В 1955 г. Д. Ариоп показал, что в изолированных хлоропластах может быть осуществлен весь процесс фотосинтеза. Важво отметить, что хлоропласты имеются пе только в клетках листа. Последнее явление (хлорофиллопосность зародыша) привлекает внимание систематикой растений. Исследования покезалв. что структура хлоропластов, расположенных в других органах растения, так же как и состав пигментов, сходны с хлоронластами листа. Зто дает основания считать, что они способны к фотосинтезу. В том случае, если они подвергаются освещению, по-видимому, в них действительно происходит фотосинтез. Так, фотосинтез хлоропластов. расположенных в остях колоса, ыожет составлять около 30% от общего фотосинтеза растения. Позеленевшие па свету корни способны к фотосинтезу. В хлоропластах, находящихся, в кожуре плода до определенного этапа его развития, также может идти фотосинтез. Согласно предполвжению А. Л. Кур-санова, хлоропласты, расположенные вблизи проводящих путей, выделяя кислород, способствуют повышен иго интенсивности обмена веществ ситовидных трубок.[ . ] Но, конечно, маловероятным является прямое сходство свойств хо-линорецептора животных клеток и мембран хлоропластов и плазматических мембран клеток.[ . ] Как уже отмечалось, под влиянием экзогенного ацетилхолина наблюдается выход в среду ионов №+ и К+ из интактных хлоропластов. Внутри хлоропластов гороха концентрация этих ионов выше, чем в цитоплазме, но оболочка пластиды непроницаема для них. Открывание ионных каналов под действием ацетилхолина, по-видимому, способствует пассивному транспорту ряда ионов по градиенту электрохимического потенциала. Однако протон в этом случае по-разному поступает в отдельные компартменты органелл.[ . ] Биомедиаторы, находящиеся в цитоплазме, могут взаимодействовать с предполагаемыми рецепторами наружной мембраны хлоропласта (других пластид), в результате чего происходят изменения Иа7К+ (ацетилхолин) и Са2+Л№ 2+ (катехоламины, серотонин) проницаемостей и включение систем вторичных посредников путем активации аденилатциклазы на внутренней стороне мембраны оболочки хлоропласта. Внутри хлоропласта ацетилхолин и биогенные амины могут связываться с рецепторами мембран тилакоидов, что также вызывает включение систем вторичных посредников (рис. 8.6 а, б).[ . ] Анатомическое строение листа приспособлено к тому, чтобы обеспечить поступление СОг к клеткам, содержащим зеленые пластиды. Наличие межклетников облегчает доступ СО2 ко всем клеткам. К эпидермису, расположенному на верхней стороне листа, примыкает палисадная паренхима, клетки которой вытянуты перпендикулярно поверхности листа. Палисадная паренхима — это основная ассимиляционная ткань листа, особенно богатая хлоропластами. Густая сеть жилок в листе пе только облегчает снабжение клеток паренхимы водой, но и способствует быстрому оттоку из листа углеводов, образующихся в процессе фотосинтеза. Для того чтобы процесс фотосинтеза протекал нормально, в клетки к зеленым пластидам должен непрерывно поступать СОг. Небольшое количество СОг образуется в самом листе в результате дыхания. Однако основным поставщиком СО2 служит атмосфера. Количество СО2 в атмосфере составляет около 0,03%. В результате поглощения листом СОг возникает градиент концентрации этого газа, что и вызывает непрерывную диффузию СОг в направлении фотосинтезирующих органов растения. СОг диффундирует из более дальних слоев атмосферы в близлежащие к листу и далее в межклетные пространства, из межклетников в клетки н далее к хлоропластам. Чем быстрее иснользуется СОг в процессе фотосинтеза, тем больше падает ее парциальное давление в межклетниках и тем быстрее поступает в них СОг. В процессе диффузии ток COj встречает сопротивление. Исследования показали, что в воде сопротивление диффузионному току СОг примерно в 1000 раз больше, чем в воздухе. Всякое перемешивание среды (воздуха или воды) способствует более быстрой диффузии СО 2 к листу. Кроме внешнего сопротивления, которое встречает С02 при диффузии до поверхности листа, существует еще внутреннее сопротивление (в самом листе).[ . ] У животных значительное количество ацетилхолина найдено во фракциях митохондрий. Была сделана попытка обнаружить ацетилхолин в хлоропластах листьев растений. Однако не у всех исследованных растений удалось обнаружить в пластидах ацетилхолин (табл. 1.2).[ . ] Наряду с минеральным питанием основой жизнедеятельности растений является усвоение углекислоты, происходящее в определенных частях клетки — пластидах (лейкопласты, хлоропласта и хромопласты).[ . ] Необходимо иметь в виду, что различные структуры, встречающиеся в незараженпой клетке, по своей морфологии могут напоминать вирусные включения. Так, в молодых хлоропластах корней фасоли найдены кристаллы запасного белка и связанные с ними пузырьки [1267]; кристаллические включения обнаружены в хлоропластах внешне здоровых листьев Масайатха [1360]. Маринос [1146] описал пластиды, содержащие вакуоли и мембрапы, в меристеме глазков клубней картофеля. Он назвал эти образования «ноли-функциопалыгами пластидами», но возможно, что некоторые из обнаруженных им ультраструктур возникли в результате «замаскированной» вирусной инфекции клубной.[ . ] Устьица располагаются большей частью на нижней, по у некоторых видов и на верхней поверхности листа. Клетки эпидермы, подобно клеткам мезофилла, несут немногочисленные крупные хлоропласты, а иногда содержат всего одну крупную зеленую пластиду. Листья некоторых обитающих в тени видов при освещении солнечным или рассеянным светом приобретают голубоватый, сине-зеленый, бронзовый или черновато-зеленый металлический оттенок, который меняется в зависимости от того, под каким углом рассматривать лист. Изменение окраски листа селагинелл связано с преломлением и дифракцией света в поверхностном слое оболочки эпидермальных клеток.[ . ] Все организмы, способные к фотосинтезу, содержат хлорофилл (немногие бактерии, способные к фотосинтезу, имеют пигменты, отличающиеся от хлорофилла). Это зеленый пигмент, содержащийся в особых органоидах растительной клетки — хлоропластах. Они взвешены в цитоплазме клетки в виде мельчайших зеленых гранул и обладают способностью движения в зависимости от освещения. В зеленых листьях высших растений хлоропласты представляют собой мелкие белково-липоидные тельца. В клетке их содержится от 20 до 100. Общее количество хлоропластов во взрослом дереве достигает десятков и сотен миллиардов, но у низших одноклеточных водорослей бывает несколько или даже одна зеленая пластида. Величина зеленой пластиды у высших растений обычно 3—5 мк.[ . ] Па свету образуется хлорофилл. Короткие тилакоиды располагаются стопкой друг над другом и образуют тилакоиды гран. Пластиды достигают окончательного размера (рис. 30). Непосредственно ив про-пластид могут образовываться и бесцветные пластиды (лейкопласты— амплопласты) (рис. 31). Лейкопласты чаще всего локализованы в клетках запасающих тканей. Во многих случаях в лейкопластах ламеллы сохраняют связь с внутренней оболочкой. В строме лейкопластов располагаются крахмальные зерня, осмио-филыше глобулы, белковые включения. Эти глобулы располагаются сплошным слоем под оболочкой пластид.[ . ] Форма лейкопластов чаще всего почти шаровидная, она изменяется лишь в тех случаях, когда в них содержатся продолговатые крахмальные зерна или выкристаллизовавшийся белок. Согласно данным Баденхоизена (1962), для формирования крахмального зерна внутри пластиды необходимы внутренние мембраны, которые в лейкопластах распределяются в мелкозернистом гомогенном матриксе, не образуя характерных ламеллярных структур, типичных для хлоропластов.[ . ] Однако результаты многих наших наблюдений не могут быть объяснены с точки ирония классической схемы распределения слоев ГЛ, 1Л1 и ЫН. Результаты недавней работы Барка и Стюарта (личное сообщение), проведенной с пластидиыми мутантами табака, позволяют предполагать, что нормальный процесс развития листа, вероятно, гораздо болео сложен и изменчив, нежели это считали ранее. Для того чтобы объяснить присутствие небольших островков нормальных клеток в палисадной ткани, мутантной по слоям Ы1 и ЫН, ученые предположили, что нормальные эпидермальные клетки (Ы) могут мигрировать в палисадный слой (Ы1) па некоторой стадии развития листа. Эти клетки (Ы) приобретают свойстиа палисадных, и таким образом возникают занявшие чужое владение островки клоток с нормальными хлоропластами. Мы предположили, что островки: темно-золепых клеток в ткани пунктирного тина образуются именно в результате такого внедрения. Ткань приобретает характерный «пунктирный» вид после того, как слой Ы1, содержащий вирус в высокой концентрации, покрывается эпидермисом Ы темно-зеленого типа. Таким образом, островки темно-зеленых клеток во взрослом листе представляют собой потомство клеток слоя Ы, мигрировавших в слой 1Л1. Микроскопические и небольшие макроскопические островки темно-зеленой ткани, появляющиеся в других участках мозаичного листа, почти всегда связаны с верхним эпидермисом. Причина этого неясна, однако Барк и Стюарт (личное сообщение) обнаружили аналогачиое распределение островков клеток мезофилла с нормальными хлоропластами в пластидных мутантах табака.[ . ] Локализация синтеза АБК в растениях еще не установлена достаточно точно, но некоторые косвенные данные указывают на то, что большая часть или, возможно, вся АБК образуется в закончивших рост зеленых листьях и плодах. Из листьев АБК передвигается в другие части растения, такие, как апексы побегов, подавляя там рост и, возможно, вызывая переход некоторого числа почек к покою (см. гл. 11). По некоторым экспериментальным данным, центрами синтеза АБК могут служить пластиды, в частности хлоропласты.[ . ] Поскольку в этом случае первый продукт карбоксилирования — ЩУК содержит четыре атома углерода, его называют «С-4» путь, в отличие от цикла Кальвина, где образуется ФГК, содержащая три атома углерода («С-3 путь). Щавелевоуксусная кислота преобразуется в яблочную кислоту. В последующем происходит реакция транскарбоксилирования, при которой СО2 снова отщепляется от органических кислот и вступает в цикл Кальвина — присоединяется к рибулезодифосфату (рис. 44). Таким образом, сущность «С-4» пути заключается в том, что реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Как и во всяком биохимическом цикле, акцепторы (ФЕП о РДФ) регенерируют, что и создает возможность его непрерывного функционирования. Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по «С-4» пути, имеются два типа хлоропластов; 1) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки; 2) мелкие гра-нальжые пластиды в клетках мезофилла листа.[ . ] ХлоропластыПластиды, как обязательный компонент растительной клетки, были описаны Левенгугов в 1676 году. Для пластидов характерна оболочка, состоящая из двух мембран. Пластиды имеют свой собственный генетический аппарат. Белок-синтезирующую систему и способны делиться внутри клетки. По пигментному составу выделяют следующие виды пластид: бесцветные (лейкопласты), хлоропласты и хромопласты. Цвет пластид обуславливается разным набором пигментов, присутствующих внутри мембран пластида. Хлоропласты имеют зеленый цвет за счет наличия на внутренних мембранах пластиды пигментов хлорофиллов. Для всех фотосинтезирующих растений преобладающим является хлорофилл-а. На самом деле, их пять – а, б, с, д, е. Отлитчаются от способности улавливать и поглощать световые волны разной длины. Однако, для всех хлорофиллов характерно поглощение с двумя максимумами – в синей и красных частях света. В хромопластах, который имеют красно-оранжевый цвет, тоже содержаться хлорофиллы, но они маскируются большим количеством пигментов каротиноидов. К каротиноидам относятся две большие группы: каротины (ненасыщенные углеводороды) и ксантофиллы (окисленные производные этих соединений, содержащие кислород в виде гидроксильных и эпоксидных групп). Функция каротиноидов – помощь в повышении эффективности фотосинтеза путем поглощения света той длины волны, которая слабо воспринимается хлорофиллом. Вторая функция – передача хлорофиллу этой дополнительной энергии. Поглощение каротиноидов осуществляется в сине-фиолетовой части спектра. При формировании растительной клетки развитие пластид как обязательных структур происходит из пропластид. пропластида представляет собой мелкий двухмембранный пузырек, диаметром от 0,4 до 1 мкм, не имеющий каких-либо внутренних особенных черт строения. Мембраны пропластиды называются внутренней и внешней. Внутренняя мембрана пропластид может давать небольшие складки или образовывать мелкие вакуоли. Особенно много пропластид в делящихся тканях растений (меристемах). Увеличение числа пропластид происходит путем деления и дальнейшая их судьба зависит от условий развития растений. Если условия связаны с наличием света, то пропластида через стадию бесцветной промежуточной структуры лейкопласта переходит в зеленую структуру – хлоропласт. Лейкопласты образуются из пропластид в темноте. Сначала происходит увеличение объема пластиды и формируется маса мелких пузырьков внутри, которые скапливаются в отдельные зоны и сложные решетчатые структуры, которые называются проламелярные тела. При освещении этих структур, эти мембранные пузырьки быстро реорганизуются и развивается полная система внутренних мембран, которые характерны для нормально организованного хлоропласта. Если же освещение не наступает и лейкопласты остаются в запасающих тканях растения, они приобретают шаровидную и элипсовидную форму. В них мало белков. Система внутренних мембран не развивается и лейкопласты, как бесцветные пластиды в запасающих тканях, выполняют функцию накопления различных веществ в виде включений. В темноте лейкопласты способны накапливать крахмал и тогда они превращаются в аминопласты. Аминопласты сплошь нафаршированы гранулами крахмала и полностью утрачивают какие-либо внутренние мембранные структуры. Если в лейкопластах накапливается в виде запасного продукта липидный компонент – жировые капли, то тогда лейкопласты превращаются в элайопласты. У лейкопластов могут накапливаться и гранулы белка и тогда это структура называется алейроновым зерном. Если лейкопласт на какой-нибудь промежуточной стадии подвергнуть искуственному отсвещению, то от внутренней мембраны лейкопласта отшнуровываются небольшие пузыречки – везикулы, которые накапливаются в строме пластиды и формируют систему тилакоидов. Граны, ламеллы, все такое. Таким образом, внутри зрелого хлоропласта есть четыре вида мембран – внешняя, внутренняя, ламеллы и тилакоиды. Хлоропласт является зеленым пластидом. Там осуществляется фотосинтез. Насыщение внутренними мембранами сильно зависит от освещенности, минерального питания, сезона года и гормонального статуса растения. Обычно хлоропласты имеют удлиненную форму, ширина их варьирует от двух до четырех микрометров, длина 5 – 10 мкм. Так же, как и у митохондрий, наружные мембраны хлоропластов обладают избирательной проницательностью и транспортируют большое количество чего-то там. А внутренние мембраны характеризуются слабой инициативой. У каждой мембраной ширина около 7 нанометров, расстояние между ними 20 – 30 нм. Внутренняя мембрана способна образовывать складчатые впячивания внутрь стромы. Обычно, ламеллы в строме лежат параллельно друг другу и не образуют связей между собой. Тилакоиды же имеют форму дисков, внутреннее пространство называется люменом. В строме хлоропласта тилакоиды могут располагаться поодиночке. Такие структуры называются хлоропластами с агранулярными тилакоидами. Число тилакоидов на одну гранулу варьирует от несокльких штук до 70 и более. Размер гран достигает примерно 0, 5 мкм, поэтому граны таки видны внутри хлоропластов. Полости камер тилакоидов всегда замкнуты и не переходят друг в друга. При естественном развитии хлоропласт в процессе жизнедеятельности растения в осенний сезон года переходит в структуру хромопласта, которая характеризуется тем, что преобладающим пигментом становится каротин. С другой стороны, бывает и развитие хромопласта минуя стадию хлоропласта. У растений, которые не подвергаются воздействию света. Морковь, например. Однако, если морковь посажена не глубоко и верхняя часть подвергается воздействию света, то могут получаться и злоропласты. На зеленой моркови есть солонин. Не нашли то, что искали? Воспользуйтесь поиском:  (пока оценок нет) (пока оценок нет) Сохранить себе в: |
